
HUMANITAT VERSUS ANIMALITAT
On rau la diferència entre un ésser humà i un animal? Som realment un animal? Som una espècie mès, una de tantes? Tan sols som més intel.ligents i prou?
 |
| La lloba Luperca i Ròmul i Rem, fundadors mítics de Roma Imatge: xavianthropus, photo CC - Localització: Museu Arqueològic de Tarragona |
HUMANS I SIMIS, QUI VE DE QUI
Un detall diferencial de la condició humana el trobem en la seva contundent inespecialització o indeterminació corporal. Entre els mamífers, els primats són també poc especialitzats, però l'èsser humà encara ho és menys. Darwin i altres pares de la teoria de l'evolució mai van donar relevància a aquest fet i des d'aleshores ençà no ha sigut explicat satisfactòriament. Podem dir que els humans mantenim tota la vida paràmetres de no-especialització i polivalència, a l'estil de les "cèl.lules mare" (salvant les distàncies) o dels casos de "neotènia" animal en que característiques juvenils o embrionàries perduren durant l'estat adult (és el cas del famós axolot mexicà, una salamandra que conserva les brànquies externes del seu estat larvari també en la fase adulta).
 |
| Neotènia. Els axolot (Ambystoma mexicanum) mantenen les brànquies, un caràcter larvàri, durant tota la seva vida adulta. Imatge: Viquipèdia, Erzengel (domini públic) |
El zoòleg alemany Julius Kolmann (1834-1918) va ser l'introductor el 1884 del concepte de neotènia en animals, i l'anatomista holandès Louis Bolk (1866-1930) va aplicar-la cap al 1890 per a explicar l'evolució humana mitjançant un retard sostingut en la nostre morfogènesi o ontogènia (desenvolupament morfològic d'un ésser viu al llarg de tota la seva vida). Bolk sosté que els humans no deixem de ser fetus que mai assolim la maduresa i parla dels humans com a "fetus de primats genèticament estabilitzats". Podem dir que comparativament amb els micos, el nadó humà neix prematur (hipòtesi evolutiva del "part prematur"), malgrat que la durada de la gestació en humans, nou mesos, és objectivament superior als vuit mesos de mitjana d'embaraç en simis. Tot i que la proliferació neuronal té lloc majoritàriament durant el desenvolupament embrionari (a partir de cèl.lules mare i cèl.lules progenitores), en humans la neurogènesi és perllonga fins l'adolescència i de manera localitzada fins l'edat adulta.
Molts evolucionistes com Stephen Jay Gould i Desmond Morris, entre d'altres, afirmen que el canvi adaptatiu clau en la evolució humana és precissament el no canvi, és a dir, un procés de neotènia o retenció generalitzada a nivell morfològic que caracteritza el cos humà, i que els simis i altres micos encara retenen en les etapes fetals o inicials del seu desenvolupament: rostre aplanat i ample, coll erecte, cervell i ulls grans, ossos del crani prims, cos i cara glabres (sense pels), nas i mandíbules petites, arc supraciliar reduït i postura erguida.
Molts evolucionistes com Stephen Jay Gould i Desmond Morris, entre d'altres, afirmen que el canvi adaptatiu clau en la evolució humana és precissament el no canvi, és a dir, un procés de neotènia o retenció generalitzada a nivell morfològic que caracteritza el cos humà, i que els simis i altres micos encara retenen en les etapes fetals o inicials del seu desenvolupament: rostre aplanat i ample, coll erecte, cervell i ulls grans, ossos del crani prims, cos i cara glabres (sense pels), nas i mandíbules petites, arc supraciliar reduït i postura erguida.
 |
| Neotènia en humans. El crani adult humà reté molt més les proporcions infantils dels homínids que no pas el crani d'un ximpanzé adult. Aixó permet que el cervell humà pugui continuar creixent durant més temps. Imatge: https://antroporama.net/category/antropologia/ |
El darwinisme ha portat molta confusió. A l’Origen de l’Home (The Descent of Man, and Selection in Relation to Sex,1871) Darwin va deixar dit que «La principal conclusió a la qual aquí s'ha arribat, ..., és que l'home descendeix d'alguna forma altament menys organitzada... i serà, em sap greu pensar-ho, molt desagradable per a molts. Però amb prou feines hi caben dubtes de que descendim de bàrbars”». Després d’això la idea popular i pseudo-científica que ha anat quallant majoritàriament des del segle XIX fins ara és que l’home prové del mico. Però en realitat no estaria clar qui ve de qui. Avui hi ha consens acadèmic sobre que simis i humans van compartir un avantpassat comú, però voler veure a partir d’aquí un origen simi dels humans no deixa de ser una interpretació esbiaixada.
 |
| Seqüència alternativa de l'evolució dels grans simis a partir d'un avantpassat comú homínid Imatge d'internet modificada amb inversió seqüèncial |
Si observem els trets morfològics dels cranis dels simis i dels primats en general, trobem que la hipermorfosi màxilo-facial (allargament del musell, creixement dels ullals, aparició de l’arc superciliar) i canvis en el neurocrani (desenvolupament de la cresta sagital), són clarament caràcters d’especialització secundària, mentre que la retenció d’un rostre pla i un neuro-crani arrodonit i gran en relació als maxilars, és una característica primària compartida pels humans, els simis nounats i els fetus dels primats en general. Primats arcaics com el tarser retenen també aquestes proporcions craneals més “primitives” en l’edat adulta.
 |
Tarser de les Filipines, Tarsius syrichta. Els tarsers
amb trets d'una primerenca evolució "humanoide" són primats molt primitius (fòssils des de l'Eocè) i amb retenció en fase adulta de caràcters juvenils no hipertrofiats: musell reduït i neurocrani arrodonit.
Imatge: Viquipèdia, Jasper Greek Golangko - copyrighted free use
|
Parlem doncs d’una “macro-evolució” explicada per processos d’heterocronia (terme encunyat per Ernst Haeckel, 1834-1919; del grec heteros: diferent i chronos: temps). Aquests processos consisteixen en alteracions cronològiques en el ritme de desenvolupament morfològic d’un organisme que provoquen transformacions en la mida i forma final d’aquest. Representen una pedra angular del procés evolutiu no explicable per la genètica de poblacions i la selecció natural, i estarien darrere dels patrons estructurals (salts morfològics en el registre fòssil, trets no adaptatius, homologies i plans corporals) que han conduït a l'aparició de l'èsser humà i altres espècies innovadores pel camí. La “microevolució” (com per exemple l’aparició de cavalls i cebres com a espècies diferenciades) tindria més a veure amb processos de tipus “darwinista” i d'abast més limitat, com la formació de races i espècies mitjançant la selecció natural i sexual, l’adaptació al medi i l’aïllament i deriva gènica.
Els processos heterocrònics ja van ser identificats per Haeckel a finals del segle XIX i estudiats sistemàticament per Walter Garstang i Gavin de Beer durant la primera meitat del segle XX, però no es van tornar a actualitzar fins que Stephen J. Gould publicà el seu llibre Ontogènia i Filogènia el 1977. Gould va analitzar en detall els processos heterocrònics que condueixen a morfologies infantils: la paedomorfosi (del grec paido: nen-, morph:forma i o-sis: procés), o avançades: la peramorfosi (del grec pera: més lluny, més enllà). La paedomorfòsi i la peramorfòsi permeten interpretar els grans canvis morfològics entre ancestres i descendents en moments crucials de l’evolució (events evolutius), sense necessitat de recórrer a l’acumulació gradual de petits canvis durant milions d’anys per explicar l’origen i la diferenciació de les espècies. Des del punt de vista de la heterocronia podem interpretar l’origen de l'èsser humà com provinent d’una línia evolutiva de fons, ancestral i paedomòrfica, caracteritzada per la retenció sostinguda i generalitzada de morfologies infantils de tipus “humanoide”. I l'origen dels simis i primats en general com a branques derivades, peramòrfiques, caracteritzades pel desenvolupament d'especialitzacions secundàries d'hipermorfosi (en major o menor grau) del tipus primat “animaloide”. Aquest segon origen seria també el cas dels nombrosos homínids extingits en el passat.
A partir de Gould va prenent força la biologia evolutiva del desenvolupament que estudia les relacions entre el desenvolupament ontogènic i l'evolució, "evo-devo", abreujat en anglès. Destaquen dues escoles dins de l'evo-devo, una més conservadora i genetista, externalista, que manté la selecció natural com la principal força creativa en els processos d'evolució; i una altra molt més innovadora i internalista, amb un enfoc morfogenètic de les causes de l'evolució al sostenir que la matèria viva és capaç d'auto-organizart-se en base a propietats biofísiques inherents als èssers vius, i que aquestes formes auto-organitzades són estabilitzades a posteriori pels circuits genètics.
A més del paleontòleg Gould, l'anatomista i zoòleg austríac Rupert Riedl fou un dels científics destacats que a partir dels anys 70 van començar a denunciar la insuficiència de l'adaptacionisme i la selecció natural per explicar els resultats de les noves investigacions morfològiques. Riedl no fou un qualsevol fora del sistema, va ser catedràtic de zoologia i ciències del mar a la Universitat de Carolina del Nord, catedràtic de zoologia a la Universitat de Viena i president honorífic de l'institut Konrad Lorenz. Junt amb aquest últim, va ser un dels fundadors de la epistemologia i psicologia evolutives. El 1968, el primer any com a catedràtic a la Universitat de Carolina del Nord, ha estat senyalat com l'any de la radicalització dels supòsits neodarwinistes de la "síntesi evolutiva moderna" (una suma de selecció darwiniana, genètica mendeliana, variació mutacional i genètica de poblacions) enfrontant-se a l'ortodòxia de l'establishment acadèmic tot desenvolupant les seves pròpies idees i patrons de l'evolució morfològica, també des del punt de vista ontogenènic i macroevolutiu. El seu llegat ha creat escola tant en l'evo-devo com en història i filosofia de la biologia (biologia teòrica).
Els processos heterocrònics ja van ser identificats per Haeckel a finals del segle XIX i estudiats sistemàticament per Walter Garstang i Gavin de Beer durant la primera meitat del segle XX, però no es van tornar a actualitzar fins que Stephen J. Gould publicà el seu llibre Ontogènia i Filogènia el 1977. Gould va analitzar en detall els processos heterocrònics que condueixen a morfologies infantils: la paedomorfosi (del grec paido: nen-, morph:forma i o-sis: procés), o avançades: la peramorfosi (del grec pera: més lluny, més enllà). La paedomorfòsi i la peramorfòsi permeten interpretar els grans canvis morfològics entre ancestres i descendents en moments crucials de l’evolució (events evolutius), sense necessitat de recórrer a l’acumulació gradual de petits canvis durant milions d’anys per explicar l’origen i la diferenciació de les espècies. Des del punt de vista de la heterocronia podem interpretar l’origen de l'èsser humà com provinent d’una línia evolutiva de fons, ancestral i paedomòrfica, caracteritzada per la retenció sostinguda i generalitzada de morfologies infantils de tipus “humanoide”. I l'origen dels simis i primats en general com a branques derivades, peramòrfiques, caracteritzades pel desenvolupament d'especialitzacions secundàries d'hipermorfosi (en major o menor grau) del tipus primat “animaloide”. Aquest segon origen seria també el cas dels nombrosos homínids extingits en el passat.
A partir de Gould va prenent força la biologia evolutiva del desenvolupament que estudia les relacions entre el desenvolupament ontogènic i l'evolució, "evo-devo", abreujat en anglès. Destaquen dues escoles dins de l'evo-devo, una més conservadora i genetista, externalista, que manté la selecció natural com la principal força creativa en els processos d'evolució; i una altra molt més innovadora i internalista, amb un enfoc morfogenètic de les causes de l'evolució al sostenir que la matèria viva és capaç d'auto-organizart-se en base a propietats biofísiques inherents als èssers vius, i que aquestes formes auto-organitzades són estabilitzades a posteriori pels circuits genètics.
A més del paleontòleg Gould, l'anatomista i zoòleg austríac Rupert Riedl fou un dels científics destacats que a partir dels anys 70 van començar a denunciar la insuficiència de l'adaptacionisme i la selecció natural per explicar els resultats de les noves investigacions morfològiques. Riedl no fou un qualsevol fora del sistema, va ser catedràtic de zoologia i ciències del mar a la Universitat de Carolina del Nord, catedràtic de zoologia a la Universitat de Viena i president honorífic de l'institut Konrad Lorenz. Junt amb aquest últim, va ser un dels fundadors de la epistemologia i psicologia evolutives. El 1968, el primer any com a catedràtic a la Universitat de Carolina del Nord, ha estat senyalat com l'any de la radicalització dels supòsits neodarwinistes de la "síntesi evolutiva moderna" (una suma de selecció darwiniana, genètica mendeliana, variació mutacional i genètica de poblacions) enfrontant-se a l'ortodòxia de l'establishment acadèmic tot desenvolupant les seves pròpies idees i patrons de l'evolució morfològica, també des del punt de vista ontogenènic i macroevolutiu. El seu llegat ha creat escola tant en l'evo-devo com en història i filosofia de la biologia (biologia teòrica).
UNA MICA D'HISTÒRIA TAXONÒMICA
Des dels temps de Darwin, una gran croada cientifista avança imparable per animalitzar i ignorar les singularitats anatòmiques, cognitives i socials humanes. Porta l’empenta d’un racionalisme materialista i reduccionista en excés que intenta explicar tota la realitat des dels supòsits (no sempre prou contrastats) de la ciència. Ara per ara s’ha de dir que la croada continua avançant triomfal i sense complexes amb genetistes quantificadors de l’ADN al capdavant. Quasi tot el món acadèmic, social, escolar i divulgatiu ha caigut als seus peus. Resisteixen però alguns biòlegs lliure-pensadors i filòsofs rebelds.
Aquests genetistes controlen tota la taxonomia actual i han eliminat les antigues families dels homínids (“Hominidae strictu sensu”, des de l’Homo sapiens fins l’Australopithecus) i dels pòngids (grans simis antropomorfs sense cua: ximpanzés, goril.les i orangutans). Actualment, fonamentant-se en un alt porcentatge de similitut gènica estrictament quantitatiu, que no qualitatiu, els primatòlegs genetistes adscriuen humans i ximpanzés a la tribu "Hominini"; a aquesta més els goril.les, a la subfamília "Homininae"; i a aquesta última més l’orangutà a la nova família "Hominidae". I a més sumen els gibons i siamangs (familia hilobàtids, petits simis antropomorfs també sense cua) per formar la superfamília "Hominoidea".
 |
"A venerable Orang-outang", caricatura
de Darwin publicada el 1871 a The Hornet, una revista satírica de l'època
Font: Viquipèdia (domini públic)
|
Aquests genetistes controlen tota la taxonomia actual i han eliminat les antigues families dels homínids (“Hominidae strictu sensu”, des de l’Homo sapiens fins l’Australopithecus) i dels pòngids (grans simis antropomorfs sense cua: ximpanzés, goril.les i orangutans). Actualment, fonamentant-se en un alt porcentatge de similitut gènica estrictament quantitatiu, que no qualitatiu, els primatòlegs genetistes adscriuen humans i ximpanzés a la tribu "Hominini"; a aquesta més els goril.les, a la subfamília "Homininae"; i a aquesta última més l’orangutà a la nova família "Hominidae". I a més sumen els gibons i siamangs (familia hilobàtids, petits simis antropomorfs també sense cua) per formar la superfamília "Hominoidea".
Així doncs, hem passat de distingir amb un mínim de seny entre simis i humans, a menystenir i ignorar les singularitats objectives que ens diferencien els uns dels altres. Abans de la forta irrupció de les tècniques d’anàlisi gènic i de la nova sistemàtica molecular, els primatòlegs i paleontòlegs diferenciaven les mones i micos amb cua dels simis antropomorfs sense cua (pòngids i hilobàtids). Ara els humans hem passat a ser també simis. Humans, simis antropomorfs, mones i micos, tots som simis! Queden fora els primats més primitius: tarsers, gàlags, ai-ais i lèmurs, que continuen sent prosimis.
Des d'un punt de vista més tradicional i assenyat qualificaríem de "micos" els primats amb cua llarga, de "mones" els primats amb cua curta (cercopitecs), de "simis inferiors" els gibons (hilobàtids), de "grans simis" els pòngids (orangutans, goril.les i ximpanzés), i de "homínids" els humans. Però corren mals temps per els naturalistes de la vella escola, doncs ara els genetistes a l’engròs, que no al detall, tenen molt més pes en els centres de decisió i de control acadèmic-universitaris. Quantifiquen el tant per cent de les seqüències coincidents de l’ADN i menystenen les petites divergències que marquen diferències decisives. Humans i ximpanzés compartiríem el 94 % de seqüències idèntiques (primer van trobar que coincidiem fins el 99% amb els ximpanzés i el 98 % amb els goril.les, però després ho han anat revisant a la baixa).
Els antropòlegs tradicionalistes fan notar que encara que genèticament siguem molt propers als ximpanzés i goril.les, hem desenvolupat (més aviat retingut) una forma corporal molt diferent (en realitat una forma fetalitzada) i una forma de vida completament nova. De fet la nostra ecologia és tan diferent de la resta dels animals, incloent-hi els ximpanzés, que pràcticament (evidentment diria) formem un regne apart, el regne humà.
Aquest és un estatus que s’ens nega, encara que des de l’escola actual revisionista no tenen problema en acceptar passar dels dos, tres, quatre o cinc regnes habituals en les antigues classificacions dels èssers vius: plantes i animals, també bacteris, protistes i fongs, fins als 12 regnes actuals d’eucariotes (organismes amb cèl.lules de nucli diferenciat), 10 regnes de bacteris i 3 regnes d’arqueobacteris, proposats ja fa vint anys. A hores d’ara desconec de quants regnes de criatures vives o extintes hauriem de parlar.
En els propers apartats veurem algunes diferències bàsiques morfològiques i cognitives entre els humans i els animals, diferències concretes que porten a dues realitats existencials completament distintes.
Des d'un punt de vista més tradicional i assenyat qualificaríem de "micos" els primats amb cua llarga, de "mones" els primats amb cua curta (cercopitecs), de "simis inferiors" els gibons (hilobàtids), de "grans simis" els pòngids (orangutans, goril.les i ximpanzés), i de "homínids" els humans. Però corren mals temps per els naturalistes de la vella escola, doncs ara els genetistes a l’engròs, que no al detall, tenen molt més pes en els centres de decisió i de control acadèmic-universitaris. Quantifiquen el tant per cent de les seqüències coincidents de l’ADN i menystenen les petites divergències que marquen diferències decisives. Humans i ximpanzés compartiríem el 94 % de seqüències idèntiques (primer van trobar que coincidiem fins el 99% amb els ximpanzés i el 98 % amb els goril.les, però després ho han anat revisant a la baixa).
 |
| Detall de l'etiqueta d'Anís del Mono, licor anisat amb origen el 1868 a una destil.leria de Badalona. La cara del mico, segons alguns, és la de Darwin. |
Els antropòlegs tradicionalistes fan notar que encara que genèticament siguem molt propers als ximpanzés i goril.les, hem desenvolupat (més aviat retingut) una forma corporal molt diferent (en realitat una forma fetalitzada) i una forma de vida completament nova. De fet la nostra ecologia és tan diferent de la resta dels animals, incloent-hi els ximpanzés, que pràcticament (evidentment diria) formem un regne apart, el regne humà.
Aquest és un estatus que s’ens nega, encara que des de l’escola actual revisionista no tenen problema en acceptar passar dels dos, tres, quatre o cinc regnes habituals en les antigues classificacions dels èssers vius: plantes i animals, també bacteris, protistes i fongs, fins als 12 regnes actuals d’eucariotes (organismes amb cèl.lules de nucli diferenciat), 10 regnes de bacteris i 3 regnes d’arqueobacteris, proposats ja fa vint anys. A hores d’ara desconec de quants regnes de criatures vives o extintes hauriem de parlar.
En els propers apartats veurem algunes diferències bàsiques morfològiques i cognitives entre els humans i els animals, diferències concretes que porten a dues realitats existencials completament distintes.
L'AUTOCONSCIÈNCIA
L'autoconsciència, en paraules del professor de filosofia Jordi Martí i Montllau, és ser conscients de la nostra consciència, ser conscients del que pensem, ..., pensar els nostres continguts de consciència o percepcions, elaborar coneixement a partir d'ells. L'autoconsciència, el pensar-se un mateix com un "jo" individual clarament separat de la resta de congèneres i de la totalitat del món que ens envolta, és una qualitat estrictament humana que no compartim amb la resta d'animals. Ens permet ser conscients d'allò que pensem i descubrir-nos a nosaltres mateixos com una realitat pensant. I ens permet copsar la nostra responsabilitat existencial com un ésser viu pensant, però alhora mortal.
L'autoconsciència tardem en assolir-la. Comencem la nostra biografia amb una consciència bastant onírica. Venim al món despreocupats i confiats, queden lluny encara la responsabilitat, la preocupació existencial i encara més la consciència de tenir un cos mortal. Quan som infants i comencem a parlar, ens referim a nosaltres mateixos amb el nostre nom propi, i no deixem de fer-ho fins que tenim entre dos i tres anys, quan comencem a autodesignar-nos com a "jo". Abans d'aixó, i no és un detall menor, hem hagut d'aprendre a caminar, és a dir, a dominar la posició vertical, erguida, pròpia dels humans. Fins aleshores ens hem desplaçat bàsicament a quatre potes. Durant el primer any de la nostra vida compartim la posició horitzontal pròpia dels animals i entre els 12 i 18 mesos començem a caminar dempeus. Psicològicament, entre els 6 i 18 mesos despertem a la "fase del mirall", descobrint la pròpia imatge, i entre els 18 i 24 mesos ens reconeixem en les fotografies. Aquesta capacitat de reconèixer-se en un mirall també la podem trobar en els simis, el grup animal més proper als humans, i en el elefants i els dofins. Sì bé demostren assolir un cert grau d'autoconsciència pròpia i del seu cos, per aquests animals arribar a l'estadi del "mirall" no equival a assolir plenament el despertar del "jo". Reconèixer la imatge reflectida és tan sols la primera fase del subtil procès humà cap a la gènesi del "jo", envers l'autoconsciència sobre la existència pròpia i la inexorable mort (la mort com a mínim del propi cos, amb això crec que hi estarem tots d'acord).
Des d'un punt de vista morfològic, algunes diferències bàsiques entre la forma i estructures humanes i la dels animals, condicionen totalment com estem i com ens relacionem amb el món, uns i altres. Aquestes diferències elementals esdevenen clau a l'hora de possibilitar el "fet diferèncial", la différence! que en dirien els francesos. Aquí però no parlarem ni de catalanitat ni de sexualitat (si em permeteu la broma), sinó d'humanitat versus animalitat.  |
| Ernst Haeckel, biòleg i naturalista alemany. Va desenvolupar la teoría de la recapitulació: l'ontogènia (desenvolupament individual) recapitula la filogènia (història evolutiva dels organismes). Autor de "La filogènia sistemàtica" (1894) i "Antropogènesi" (1874). Fou més partidari de la heterocronia i del transformisme lamarckià de les espècies que de la selecció natural darwiniana. Imatge del 1906: Viquipèdia (domini públic) |
L'autoconsciència tardem en assolir-la. Comencem la nostra biografia amb una consciència bastant onírica. Venim al món despreocupats i confiats, queden lluny encara la responsabilitat, la preocupació existencial i encara més la consciència de tenir un cos mortal. Quan som infants i comencem a parlar, ens referim a nosaltres mateixos amb el nostre nom propi, i no deixem de fer-ho fins que tenim entre dos i tres anys, quan comencem a autodesignar-nos com a "jo". Abans d'aixó, i no és un detall menor, hem hagut d'aprendre a caminar, és a dir, a dominar la posició vertical, erguida, pròpia dels humans. Fins aleshores ens hem desplaçat bàsicament a quatre potes. Durant el primer any de la nostra vida compartim la posició horitzontal pròpia dels animals i entre els 12 i 18 mesos començem a caminar dempeus. Psicològicament, entre els 6 i 18 mesos despertem a la "fase del mirall", descobrint la pròpia imatge, i entre els 18 i 24 mesos ens reconeixem en les fotografies. Aquesta capacitat de reconèixer-se en un mirall també la podem trobar en els simis, el grup animal més proper als humans, i en el elefants i els dofins. Sì bé demostren assolir un cert grau d'autoconsciència pròpia i del seu cos, per aquests animals arribar a l'estadi del "mirall" no equival a assolir plenament el despertar del "jo". Reconèixer la imatge reflectida és tan sols la primera fase del subtil procès humà cap a la gènesi del "jo", envers l'autoconsciència sobre la existència pròpia i la inexorable mort (la mort com a mínim del propi cos, amb això crec que hi estarem tots d'acord).
QUESTIÓ DE VERTICALITAT
 |
| Boiximà del Kalahari |
Un d'aquests detalls fonamentals és la situació del forat magne (foramen magnum) a la base del crani. En els humans, aquest forat on s'engalza la columna vertebral amb el crani, es situa en la part inferior central del mateix, permetent així una posició erguida del tronc i la marxa bípeda. D'aquesta manera, els nostres músculs del coll no necessiten ser tan forts com en el cas dels grans simis homínids (orangutans, goril.les i ximpanzés) per mantenir dret el cap. Podem dir literalment que no ens pesa el cap! I això que de mitjana, en posició dreta o vertical, portem més de 5,5 kilos recolzats sobre les cervicals (i si hi dobleguem el coll cap endavant, aquest pes es pot arribar a multiplicar per cinc). Aquesta posició inferior central del gran forat occipital del crani humà determina que el nostre grau de confort com a espècie, a l'hora de caminar amb el cap dret sobre les extremitats inferiors, sigui superior al dels simis. En aquests animals, els més propers anatòmicament, el foramen magnum es troba un tant desplaçat cap a la part posterior del crani, i en la resta d'animals el desplaçament encara és més pronunciat i evident. En tant que el cap humà resta en equilibri vertical sobre el tors, en la resta de mamífers el cap es manté per davant del coll, és a dir, en un desequilibri permanent. Certes mones com els macacos són bastant bípedes en l’etapa juvenil primerenca, aptitud que perden totalment a l’arribar a l’adolescència (Michejda i Lamey, 1971). Alguns investigadors veuen una connexió directa entre aquest canvi de conducta i el retrocés del foramen magnum cap a la part posterior del cap durant el creixement (Masters i altres, 1991). Entre els primats adults, els gibons són el únics que també poden "caminar" amb certa naturalitat desplaçant-se amb les extremitats posteriors, encara que la seva anatomia esta especialment modificada per practicar la locomoció per braquiació: avancen pels arbres grontxant-se d'una branca a l'altre amb uns braços que fan quasi el doble que les cames. I entre els lèmurs (un primat prosimi), el sifaka està especialment adaptat per desplaçar-se saltant en posició vertical, recordant una mica als cangurs.
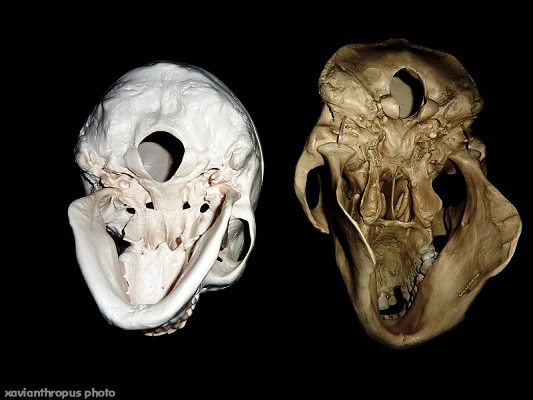 |
| Models de crani humà i d'orangutà, vista inferior. Observeu el desplaçament cap enrere del foramen magnum en el crani de l'orangutà (dreta) comparat amb la posició central del foramen magnum en el crani humà (esquerra) |
Els humans adoptem la verticalitat com una manera d'estar en el món, a diferència dels animals, que cauen en major o menor grau en la horitzontalitat, i acostumen a desplaçar-se amb el seu eix o columna vertebral en paral.lel al terra, ja siguin animals terrestres, aquàtics o aèris. Dins dels vertebrats, els quadrúpedes ("quatre peus") es desplacen caminant o corrent amb les quatre extremitats, les serps avancen amb ondulacions laterals del cos, els ocells i ratpenats volen amb les ales (les seves extremitats davanteres), els peixos nedan impulsant-se amb les seves aletes i flexions laterals del cos, i els cetàcis ho fan amb moviments verticals de la aleta caudal horitzontal (en els peixos és vertical) ajudats per les extremitats anteriors convertides en aletes pectorals. I entre els invertebrats podem citar com exemple representatiu els insectes i les seves innumerables espècies caminadores, saltadores, voladores, nedadores, ... totes elles sempre desplaçant el seu cos en un pla horitzontal, tant la formiga al caminar com la papallona o l'abella al volar.
En aquest sentit, solament els éssers humans i les plantes podem dir que es manifesten morfològicament a partir d'un eix vertical que va de dalt a baix o de baix a dalt, segons es miri. En el humans el pol neuro-sensorial (el cap) s'orienta cap al cel i en els vegetals les arrels "sensorials" s'enfonsen en la terra (geotropisme) tot buscant els nutrients i l'aigua. En canvi el pol metabòlic-reproductor es posiciona inferiorment en els humans i superiorment en les plantes (flors i fruits). La part mitja respiratòri-circulatòria ocupa la zona central en ambdós casos, encara que també invertida. La tràquea humana "baixa" des de dalt i el tronc de l'arbre (o tija d'una planta) creix cap amunt, el mateix respecte dels bronquis i les branques. Bronquiols i branquillons ja es reparteixen més lliurement a l'igual que els sacs alveolars i les fulles.
En canvi, en els animals, subjugats d'alguna manera per les forçes gravitatòries terrestres, l'eix corporal es manté majoritàriament sempre horitzontal al terra. N'hi ha, però, excepcions. Alguns grups d'animals participen fins a cert punt de l'impuls evolutiu ascendent que du a estar dempeus sobre la terra. És el cas d'alguns marsupials com els cangurs, d'aus com els pingüins i també de peixos com els cavallets de mar. Els extints dinosaures serien un altre cas d'aproximació barroera vers la posició erguida.
NEUROGÈNESI
La mielina, la "substància blanca" lipídica que recobreix els axons, part basal de les neurones, hi juga un paper destacat en aquest desenvolupament nerviós. Permet accelerar les conexions neuronals mitjantçant enllaços químic-elèctrics, les sinapsis. Gràcies a elles es generen multitud de circuits inter-neuronals durant el període embrionari i els primers dos anys de vida, com ara la xarxa interneuronal descoberta per G. Lanuza en la medul.la espinal i que permet l'alternança coordinada de les dues extremitats inferiors al caminar.
Després dels dos anys i fins l'adolescència, apareix el fenòmen de la "poda sinàptica" o destrucció de les connexions sobreres per tal d'optimitzar l'eficiència de la xarxa neuronal restant. En aquest procès d'autodestrucció s'arriben a perdre fins a la meitat de les sinapsis generades en les etapes prenatal i postnatal primerenca .
Investigadors com Miller i Rilling van descobrir un retard notable, passada l'etapa adolescent en humans, de la mielinització definitiva de totes les àrees associatives del neocòrtex, la part més evolucionada de "l'escorça cerebral" o còrtex (desenvolupat especialment en primats i humans). Aquesta part del cervell té a veure, entre d'altres, amb funcions cognitives superiors que van des de les interaccions socials, autoreflexió i resolució de problemes fins al llenguatge humà). S'ha vist que aquest retard en la mielinització del neocòrtex es manté en humans aproximadament fins als 30 anys, un fet excepcional que no es dona en cap primat. Abans d'aquest descobriment se sabia que el creixement primerenc ràpid del cervell humà s'allarga fins al 6 anys i no s'assoleix el ple desenvolupament fins als 23 anys aproximadament, mentre que en els ximpanzés aquest desenvolupament s'atura just a l'arribar al primer any de vida.
Els micos, de mitjana, neixen amb un 70 % assolit del tamany del cervell adult. Els petits simis com el gibó ho fans amb el 64 %, els goril.les amb el 56 % i els altres grans simis com orangutans i ximpanzés amb el 38 i 36 %, respectivament. Els humans ho fem amb un 26 %. Dit d'altra manera, la grandària del cervell humà nounat equival a una quarta part d'un cervell adult, mentre que la massa cerebral del ximpanzé nounat (l'espècie més propera als humans genèticament parlant) és aproximadament un terç de la dels adults. Concretament les proporcions en humans són de 335g en nounat per 1300 g en adults. En els ximpanzés les xifres són de 128 g en nounat per 360 g en adult.
Actualment, però, hi ha consens en que la diferència bàsica entre un cervell de simi i un d'humà no és tant una questió quantitativa de més o menys massa cerebral, sinó una questió de diferència qualitativa, deguda als canvis tardans en el cervell humà que li permeten una major plasticitat. Aquest període estès de desenvolupament cerebral dóna als humans l'oportunitat d'una maduració cerebral més gran durant l'adolescència i fins el començament de l'edat adulta, sempre en funció de l'entorn. A destacar l'adolescència com el període clau per l'adquisició d'habilitats socials i formes adultes de llenguatge i comunicació.
DENTICIÓ
Una altra manifestació molt il.lustradora d'aquesta no-especialització humana la trobem en les nostres dents. Anatòmicament són el més dur de tot el cos, més que els ossos de l'esquelet. En concret l'esmalt dental és el teixit més dur de l'organisme, amb un 95 % de sals de calci i fòsfor en forma de cristalls exagonals de hidròxiapatita (un fosfat de calci), un 2 % matèria orgànica i un 3 % d'aigua. La dentina per sota de l'esmalt conté un 70 % d'aquestes sals, al igual que la matriu extracel.lular dels ossos, formada també per hidròxiapatita. L'esmalt format completament està tan mineralitzat que és incapaç de contenir cèl.lules vives entre els cristalls d'apatita que s'agrupen en forma de prismes recobrint la dentina.
Podriem dir que en exemples anatòmics tan durs com les dents, inserides alhora en els durs ossos maxil.lars del crani, trobem una excel.lent imatge cristalitzada que evidencia el fort impuls de retenció en humans enfront de l'especialització unilateral animal. Altres processos de cristalització sensible* en laboratori són força coneguts pels agricultors i ramaders biodinàmics. Es fan servir per veure les propietats qualitatives vitals de qualsevol substància orgànica, mitjançant un procès estandaritzat d'assecament i cristalització de dilucions salines, en aquest cas clorur de coure i no hidròxiapatita, mesclades amb la substància a estudiar. Las seves imatges no expressen reaccions químiques sinó la materialització de camps de força.
Les tendències que els vertebrats superiors i la resta d'animals desenvolupen unilateralment i de forma preeminent les unes sobre les altres, són expressió de la naturalesa de les seves forces formadores morfogenètiques, forces grupals animals (d'ànima, animades) que intervenen en la zoogènesi de cada espècie i que queden fixades posteriorment en el seu patrimoni gènic (gene pool). En aquestes forces morfogenètiques animals la manifestació del "jo" queda relegada a nivell grupal d'espècie, al contrari que en l'antropogènesi, on les forces del "jo" impregnen directament la formació del cos humà.
 |
| Humans i gossos. Compareu el rostre pla de la noia, la uniformitat de les seves dents i l'absència d'espais entre elles amb el rostre del gos "distorsionat" pel creixement del musell, la hipertròfia dels canins i l'aparició d'espais interdentals. |
Tots sabem que les nostres dents no creixen fins a convertir-se en grans molars, grans canins o grans incissius, és a dir, no cauen en la hipertròfia unilateral que els diferents tipus de mamífers sí que mostren. O no desapareixen per ser substituïdes per un bec corni, com és el cas de les aus actuals (l'arqueòpterix, primer ocell conegut, tenia dents).
*Cristalització Sensible: https://www.terradinamica.cat/metodologia/
UNILATERALITAT ANIMAL
REMUGANTS
D'acord amb el tipus de dents més desenvolupades o la seva absència, podem observar tres grans tendències en els vertebrats superiors.
En els mamífers remugants trobem la màxima expressió d'hipertròfia molar. Aquests presenten queixals de considerables dimensions adaptats a la seva dieta herbívora i d'animals remugants que pasturen i masteguen més tard. Són animals molt metabòlics i el seu aparell digestiu és un dels més complexes entre els mamífers. Tenen quatre cavitats estomacals: rumen, reticle, omàsum i quall!
 |
| Crani d'ovella (Ovis aries) latxa. Aquestes raça basca de xais sovint presenta banyes en ambdós sexes, com els bovins. Uns i altres, bens i bous, i també altres remugadors, no tenen canins ni incisius superiors. |
La vaca, el remugant arquetípic, presenta una dentadura a base de molars i pre-molars mancada dels canins i dels incissius superiors. La tipologia de les seves dents mostra un animal amb un desenvolupament exagerat del pol metabòlic-digestiu en detriment d'altres parts del cos com ara el pol neuro-cognitiu.
 |
| Vaca (Bos primigenius taurus) remugant al Matagalls |
El mot brau, que fa referència al mascle de la vaca o bou no castrat, també és utilitzat com a sinònim d'animal fer i salvatge, i és tota una icona de les forces testosteròniques cegues encara no transformades per la llum del pensament i la reflexió.
 |
| Vaques i bous braus pasturant al Delta de l'Ebre |
ROSEGADORS
En canvi, en els rosegadors observem un desenvolupament màxim dels incissius (les dents "nervioses") i uns molars comparativament esquifits. Poden presentar pre-molars però com en els remugants falten també les dents canines o intermitges. Aquest grup mostra un comportament certament inquiet. Roseguen continuament amb els incissius per compensar el creixement continu d'aquestes dents. També tenen una tendència a alliberar els membres anteriors, a seure erguits i a dur-se les "mans" a tocar de la boca per facil.litar el rosec: l'esquirol rosegant una pinya n'és un clar exemple.
 |
| Esquirol, Esciurus vulgaris. Serra de Guara, Osca |
Però com més gran és un rosegador, més es ralentitza el seu metabolisme i més recorda el tipus de mamífer rumiant. Pensem en el capibara, el rosegador sudamericà més gran que existeix (50-65 kg), o en el coipú o rata-llúdriga (fins a 10 kg), oriünd de la Patagònia però en expansió a les comarques gironines on ha arribat des de França. I també hi ha espècies de rosegadors bastant predadores u omnívores, com ara els lirons.
 |
| Coipú, Myocastor coypus. Dreta, exemplar de coipú a França, Philippe Amelant, imatge CC - Viquipèdia Esquerra, crani de coipú. Observeu el gran espai interdental, l'absència de canins i els potents incisius ataronjats. |
INSECTÍVORS
Sense ser pròpiament rosegadors, les musaranyes, petits animalons insectívors similars als ratolins, mostren un comportament encara més nerviós. Amb un metabolisme molt accelerat (el seu cor batega a 25 pulsacions per segon) són els mamífers més actius i uns dels més petits que hi ha. Tenen un llarg musell amb bigotis molt sensibles que els permeten detectar els insectes i cucs dels que s'alimente sense treva, doncs han de menjar l'equivalent del seu propi pes diàriament.
I també tenim, dins del grup dels quiròpters, el rat-penat o rata-pinyada, una mena de ratolí volador que ha anat mès enllà que qualsevol mamífer, superant de llarg la capacitat planejadora dels esquirols voladors. Els ratpenats han conquerit el mitjà aèri com si fossin ocells, però sense ser-ho. Volen, o més aviat revolotegen quan es fa fosc, gràcies a una gran membrana dèrmica o patagi que uneix uns dits extraordiàriament llargs. Són els pseudo-ocells de la nit. Animals molt sensibles i porucs, durant el dia s'amaguen de la llum i busquen refugi en coves i en construccions humanes (quant més ruïnoses i tranquil.es millor). Són animals molt sensorials que han desenvolupat unes grans orelles i un parell adicional de més petites, anomenades tragus, relacionades amb un sistema sensorial molt sofisticat, l'ecolocalització, mitjançant l'anàlisi dels ecos dels seus crits ultrasònics. És així que es poden orientar en la foscor, evitar obstacles o capturar les seves preses. "Veuen" amb les orelles! encara que no són totalment cecs. Amb prou llum poden veure en blanc i negre amb els seus reduïts ulls.
OCELLS
Els ocells encara donen un pas més que els rosegadors, esquirols voladors i ratpenats. Volen amb mestria amb les plomes remeres de les dues ales i les timoneres de la cua. Han desenvolupat a l'extrem anterior del seu cos un bec còrni que recobreix i va més enllà de la mandíbula òssia no dentada. Han canviat les dents per un bec! Els ocells no masteguen el menjar sinó que se l'empassen. Tenen un pap per emmagatzemar-ho i un pedrer amb pedretes que s'empassen per a moldre, a falta de dents, els aliments. Presenten doncs una digestió ràpida adaptada al vol. No s'entretenen gens en mastegar o remugar i tampoc separen molt els residus, excreten la femta i l'orina (àcid úric en lloc d'urea) tot junt en forma de residu semisòlid i a travès d'una única obertura, la cloaca. No tenen ni bufeta urinària ni conducte uretral.
 |
| Merla, Turdus merula. Dreta, mascle adult. Esquerra, crani de merla. Observeu el gran tamany relatiu de l'òrbita ocular i de l'anell escleròtic. |
Comparativament el sistema nerviós és gran en proporció a la mida de l'ocell. La part més desenvolupada del cervell té a veure amb les funcions relacionades amb el vol, el cerebel amb la coordinació del moviment i el telencèfal amb la navegació i la construcció dels nius. A més, els ulls dels ocells són sensibles a la llum ultraviolada, invisible pels humans, i també al verd, vermell i blau, són doncs tetracromats. La majoria de mamífers en canvi són dicromats, com els gosos, i tenen una visió en color equivalent al daltonisme vermell-verd dels humans. També és notòria la intel.ligència de còrvids i lloros, la capacitat de l'aufrany per utilitzar eines (agafa pedres amb el bec per obrir i menjar-se els ous d'estruç), la bellesa del cant del rossinyol o la destresa dels ocells teixidors fent els nius.
 |
| Àliga daurada, Aquila chrysaetos Esquerra: font Viquipèdia, Richard Bartz CC BY-SA 2.5 - Dreta: font Wikipedia, Juan Lacruz CC BY 3.0 |
Trobem doncs en els ocells que el pol neuro-sensorial tendeix a desenvolupar-se força (més que en el cas dels mamífers rosegadors, que també, penseu en les obres d'enginyeria dels castors, per exemple) en detriment d'un sistema metabòlic més "simplificat". Podriem pensar en l'àliga com l'arquetip d'aquests tipus d'animals. Vista poderosa, mirada penetrant, port majestuós, capacitat d'enlairar-se a grans alçades lluny dels lligams terrenals i lliure de la pesantor gravitatòria, ... són atributs que els humans percebem en l'aliga.
CARNÍVORS
I finalment tenim un altre tipus de mamífers en els que observem una hipertròfia clara de les dents canines, els ullals! Parlem, és clar, dels carnívors, com ara un gos o un gat domèstics, per exemple, o d'espècies salvatges com el llop i el tigre.
També tenen ullals altres mamífers poc especialitzats i una mica omnívors com el senglar i els porc. En canvi, els grans "ullals" del elefant en realitat no són canins sinó incissius. Els paquiderms són grans herbívors molt "metabòlics" però alhora tenen un pol "neuro-sensorial" molt desenvolupat. Tenen fama d'una gran memòria i intel.ligència i estàn dotats d'una trompa molt sensible.
Tornant als animals amb grans canins, tenim en els fèlids el tipus més especialitzat. Mantenen les dents incissives i alguns molars, però sobretot desenvolupen els grans ullals que els caracteritzen. Seria el cas de tigres i lleons o dels espectaculars i extints dents de sabre com el Smilodon. En el lleó trobariem l'arquetipus del mamífer carnívor predador: grans dents intermitges -els ullals- i clar predomini de la zona central del tòrax i costelles. La zona abdominal resta bastant continguda i els intestins no són molt llargs. La digestió és intensa però no extensa. Vist per davant un lleó impressiona amb els seus formidables ullals i la magnífica melena que li cobreix el cap i el pit, una amplia caixa de ressonància per el seu impressionant rugit. En canvi, vist d'esquena, podriem dir que decepciona una mica i no sembla gran cosa. No desenvolupa gaire l'extrem posterior -pol metabòlic- del seu cos. Ullals, melena, veu poderosa i urpes són el seu fort. I resulten prou convincents per fer del lleó un dels animals més representats en l'heràldica (juntament amb l'àliga i el drac mitològic).
Tradicionalment s'associa el lleó amb el coratge i la valentia, simbolitzats pel seu cor (recordeu en Ricard Cor de Lleó, valent combatent, lider guerrer i rei d'Anglaterra). El cor, a la fi, pertany a la part central del cos i equilibra amb les seves pulsacions rítmiques els extrems cefàlic i abdominal. Cor i pulmons es coordinen dins del sistema sanguini per regular els ritmes cardio-respiratòris. Amb el ritme dels batecs del cor i de les respiracions, la regió toràcica armonitza les polaritats en un organisme sa. Quan el cor perd el ritme es presenta l'arrítmia, un transtorn cardíac que pot arribar a ser greu.
INESPECIALITZACIÓ HUMANA
En els mamífers i els humans la regió toràcica està clarament separada de la regió abdominal pel múscul del diafragma. Parlem així d'una frontera objectiva, que no subjectiva, entre regió abdominal i regió toràcica. La regió cefàlica, el cap, resta unida i alhora separada del tòrax pel coll. Ens trobem doncs, davant d'una clara tripartició estructural en tres grans àrees orgàniques d'influència. En els animals, les fortes descompensacions unilaterals respecte al desenvolupament relatiu de les àrees cefàlica, tòracica i abdominal queden reflectides en la seva dentició o bec. A grans trets, podem distingir en els vertebrats superiors (ocells i mamífers), i sense menystenir tots els casos intermedis menys especialitzats, tres grans impulsos o tipus d'especialització a partir de les seves dents o becs. Així, en els ocells i els mamífers rosegadors predominarien les forçes neuro-cefàliques, amb desenvolupament de becs o dents incissives. En els mamífers remugants i altres grans herbívors les forces digestives i abdominals, amb els premolars i molars ben desenvolupats. I en els carnívors les forces toràciques cardio-respiratòries, tots ells amb bones dents intermèdies o canines i amb els fèlids com a punta de llança. El lleó, la vaca i l'àliga serien els animals més representatius de cada grup.
Arribats a aquest punt, afegir que, en els humans, trobem també un ésser tripartit però equil.librat, mancat de l'especialització animal i, per tant, "lliure" gràcies a la seva indeterminació, polivalència i totipotencialitat. Estariem davant d'un ésser equilibrat i polivalent, no especialitzat. No animalitzat a la fi.
ROSTRE HUMÀ I MUSELL ANIMAL
Els humans tenim, en comparació amb els animals, una cavitat oral excepcionalment curta. No desenvolupem un musell o boca prominent com els mamífers o els rèptils, o un bec com les aus i les tortugues. Totes les etapes fetals del mamífers presenten boques curtes, però en els humans aquesta característica és manté sempre, mentre que en els mamífers es va perdent poc o molt durant el creixement i maduració.
El rostre ample i aplanat és un altre fenòmen de fetalització o retardació propi dels humans. En els primats inferiors ja trobem la tendència a reduïr el musell, amb un paladar un 30 % més curt que el dels mamífers insectívors. En els micos la reducció augmenta de mitjana fins al 40 % i en els humans fins al 65 %. En el cas de l'arc dental succeeix quelcom de similar. En els primats, respecte dels insectívors i micos inferiors, la seva longitud minva un 25 % i en humans un 65 %. Aquesta tendència cap a la reducció de l'arc dental va acompanyada de una reducció en el nombre de dents. Evolutivament la quantitat de dents en els mamífers tendeix a disminuir en general, tret d'algunes especialitzacions concretes. Mamífers antics prehistòrics mostren al voltant de 60 dents i els rèptils encara més. En simis i humans s'ha reduït significativament la longitud de la fila dental dels molars i pre-molars. En humans adults les dents són trenta-dos: 4 incissius, 2 canins, 4 premolars i 6 molars (comptant els queixals del seny, de tardana aparició) a cada mandíbula. En la dentició de llet falten els molars.
Quant a la forma de l'arc dental humà és molt diferent de la dels micos adults. En els primers es manté sempre curva, però en els segons arriba a ser casi rectangular. Primats inferiors com el tarser filipí i petits simis com els gibons mantenen però tota la vida un rostre més aplanat i humanoide.
En canvi, els simis antropomorfes han abandonat les característiques primitives i prototípiques d'un crani facial retingut, sense musell, sobretot els mascles adults que desenvolupen ullals espectaculars. Les femelles en canvi mantenen més les característiques fetals i juvenils amb unes dents més contingudes i un arc dental menys rectangular.
En l'evolució ascendent d'un arquetipus homínid moderat i retingut sembla que la questió de gènere és important. Els mascles cauen més en l'animalització i atenent al seu desenvolupament ontogènic individual podriem dir que la seva involució és notoriament major. Si un goril.la (educat amb una tutora humana) va ser capaç d'aprendre a comunicar-se amb els humans mitjançant llenguatge de signes, aquesta va ser Koko, un goril.la femella, esclar. Va arribar a dominar fins a 1000 paraules en el llenguatge de signes americà (ASL) i a comprende fins a 2000 en anglès. Un espectacular aprenentatge cognitiu per a l'espècie dels goril.les. També hauriem d'admetre en els mascles humans un cert handicap social evolutiu de caire testosterònic que contrasta amb la plasticitat i flexibilitat adaptatives femenina i que confirmaria alló de que "la dona és el futur de l'home".
LES DENTS I LA PARLA
L'estructura de la mandíbula i les dents humanes va més enllà de la pura funció masticatòria i alimenticia. La dentició no correspon amb cap dels patrons habituals dentals dels diferents tipus de mamífers com carnívors, herbívors, rosegadors, insectívors o fins i tot omnívors com els porcs, doncs la boca humana a més de formar part de l'aparell digestiu és alhora una estructura anatòmica que possibilita la parla juntament amb la laringe, l'òrgan propi de la fonació.
Degut a la uniformitat d'alçada i amplada de totes les dents i sobre tot a l'orientació vertical dels incisius i la falta d'espais entre les dents frontals, la dentadura humana forma davant de la cavitat oral una tanca continua que és essencial per a la formació de consonants.
DESENVOLUPAMENT DEL CRANI
Una cosa semblant succeeix amb el desenvolupament de la resta del cap. El crani d'un mico neonat s'assembla molt en proporció i forma al d'un humà. Simis i humans encara mantenen aquesta semblança en les etapes juvenils del desenvolupament, però en les fases posteriors solament els humans són capaços de mantenir les proporcions d'aquesta forma cranial comú inicial. El bonobo o ximpanzé pigmeu, per alguns autors una variació neotènica del ximpanzé comú, manté en la fase adulta una menor desviació del patrò de crani humanoide que l'altre ximpanzé, i també mostra un menor desplaçament posterior del foramen magnum. Es a dir, és manté relativament més proper al tipus universal humanoide en desviar-se menys de la seva figura juvenil.
En els micos, la forma original del crani fetal i juvenil canvia de més esfèrica a allargada, degut al creixement posterior de les mandíbules. En canvi, el neurocràni deixa de crèixer ben aviat, de manera que el creixement de la cavitat cerebral s'atura deseguida mentre continua el creixement de l'aparell masticador.
En els humans el crani tarda en ossificar-se definitivament. Neixem amb fontane.les (espais tous) i separacions entre el ossos cràneals. Aixó permet la superposició d'aquests ossos durant el naixement, degut a un crani massa gran a l'hora de passar pel canal del part. Les fontanel.les resten sense tancar després de néixer encara durant un cert temps (uns 18 mesos tarda en ossificar la fontanela anterior o major). Els ossos cranials que envolten al cervell continuen creixent fins els sis anys, amb el 90 % assolit del desenvolupament, i la resta en arribar a la pubertat. En canvi, les sutures fibroses entre els ossos del crani no ossifiquen fins a edat avançada i en alguns individus mai, com és el cas de la sutura frontal o metòpica. El desenvolupament del crani facial també s'allarga en comparació al del neurocrani. Al sis anys ha arribat al 80 % del seu creixement, però continua lentament en la pubertat i més enllà.
L'ASSUMPTE DEL MENTÓ
La mandíbula, propiament dita, o mandíbula inferior, queix o maixella, no presenta ossos premaxil.lars com la mandíbula superior. Està formada per la unió de les dos hemimandíbules o hemiarcades. En els infants humans els dos maxil.lars inferiors es mostren separats durants un temps per la símfisi o sutura mandibular, però es fusionen entre el primer i segon any de vida mitjançant l'ossificació dels teixits connectius de la símfisi. En molts altres animals pot romandre sempre sense ossificar, com per exemple en gossos i vaques, ambdós amb fort creixement frontal del crani facial.
També obsevem un desfase entre un major creixement frontal de la mandíbula i un creixement menor de l'arc dental humà. Això dóna origen a l'aparició del mentó o barbeta, un tret exclusiu nostre que no compartim amb cap animal. La mandíbula humana continua creixent una mica durant el segon septeni, quan el creixement de l'arc dental ja s'ha aturat abans coincidint amb el canvi de la dentició de llet. D'aquí el mentó lleugerament prominent.
En els simis antropoides això no passa, ja que l'arc dental continua creixent a un ritme major que la base mandibular, que també creix però menys i per tant queda sempre una mica endarrerida i fonedissa. En els simis i els altres mamífers observem també un desfase temporal en el patró de creixement de la mandíbula i de l'arc dental, però en sentit totalment contrari.
EL MISTERI DE L'OSSA INCISIVA
Els vertebrats, des dels peixos ossis fins els ocells amb bec o els mamífers amb morro, tenen dos petits ossos situats al davant de la mandíbula superior, són els premaxil.lars i són part i causa important de la prominència del musell. Envolten l'obertura nasal i part del pont del nas i baixen fins a la boca, on conformen l'àrea frontal de l'arc dental superior i allotjen les dents incissives, d'aquí el nom d'ossa incisiva. En el musell dels mamífers trobem aquests ossos separant els dos maxil.lars superiors, tot impedint-los soldar entre si els dos extrems frontals i en consequència sense cap fre que aturi el seu creixement cap endavant.
Abans de les investigacions de Goethe (1780) es creia que l'ossa incisiva era inexistent en humans. Contrariat amb aquesta afirmació, Goethe va descobrir finalment en alguns cranis humans restes de la sutura entre el maxil.lar i l'os incisivum, fet que confirmava l'existència dels premaxil.lars en humans. Aquesta sutura es pot trobar en una quarta part dels cranis d'adults humans i en la majoria dels juvenils. Sempre es troba a la banda del paladar, formant un cercle tancat per darrera dels incisius, i mai a la part externa del crani facial, on queda ja engolida pels maxil.lars al final de l'etapa embrionària.
La falta de musell en els humans, per tant, va associada a aquest ràpid engoliment dels ossos premaxil.lars per part dels maxil.lars superiors durant el desenvolupamernt primerenc, cap al final del tercer mes de gestació.
Goethe ja va veure en aquesta ràpida incorporació dels premaxil.lars una expressió de la resistència humana al desenvolupament del musell. Considerava que un dels propòsits d'aquesta resistència era la preservació d'una fila de dents tancada (detall important per a la parla). Molts mamífers presenten diastemes o espais entre les dents, com ara els remugants i els rosegadors entre els molars i els incisius, i els primats també mostren uns espais per a acomodar les parts superiors de cadascun dels grans ullals en el maxil.lar contrari pertinent. En els infants humans trobem també habitualment aquests anomenats "espais primat" i altres diastemes de creixement en la dentició temporal o de llet, un fet que facilita l'encabiment de les futures peces dentàries definitives de major tamany.
Respecte a la sutura incisiva, a més de vestigis en el paladar, en els grans simis trobem també restes d'ella a la banda exterior del crani, ni que sigui temporalment. Bolk va estudiar el tema en 1912 i va trobar que en els ximpanzés la sutura externa pot desaparèixer ja abans de néixer, en els orangutans es perd durant l'infància i més tard en els gori.les. En canvi perduren en els adults de les mones aranya.
La tendència a l'escurçament de l'os incisivum i del musell, tot mantenint un cap arrodonit, ja la trobem en primats primitius com els tarsers (amb fòssils des de l'Eocé, a principis del període Terciari), encara que hi ha importants retrocessos en etapes més tardanes de la evolució dels primats, com en els papions, gelades, drils i mandrils (tots ells amb un musell força prominent i ullals impressionants en els mascles). Aquesta tendència cap a la supressió del musell es manté però com a característica evolutiva del primats, i en els simis superiors s'observa clarament un tancament ràpid del premaxilars, encara que no aconsegueixen frenar del tot el desenvolupament del musell. Solament els humans assolim la supressió completa, obrint el camí cap a la parla. Un camí fressat de confluències sinèrgiques com les descrites fins aquí i d'altres no comentades encara.
LA COMPOSICIÓ SINÈRGICA
Altres característiques humanes són, per exemple, l'independència del ritme respiratori, al no estar condicionat per una locomoció quadrúpeda (una respiració per cada pas o gambada, o bé la independència respiratòria respecte de la termoregulació per ventilació pulmonar (el panteix dels gossos). Amdós circumstàncies faciliten la parla. També la llengua que surt des de més avall i enrera, formant la paret anterior mòbil de la faringe (molt important per modular els sons) i una laringe o garganxó situada en els adults humans molt més avall que en els mamífers (podem respirar independentment per la boca o el nas), i en angle recte respecte de la cavitat bucal (aquesta posició permet un inventari molt ample de sons). Les cordes o plecs vocals, a més, són de constitució muscular en els humans i membranoses en els primats inferiors, mentre que en els simis antropoides són d'una constitució intermèdia.
Al cap i a la fi tot un ventall de processos sinèrgics convergents per a possibilitar la parla humana.
Les diferents consequències del retard generalitzat en el desenvolupament humà respecte de la forma prototípica mamífera o primat, i fins i tot vertebrada o cordada, es junten armoniosament en el cos humà tot formant una composició sinèrgica, un conjunt potencialment molt superior a la simple suma de les seves parts.
L'objectiu del retard constant i creixent dels primats al llarg de l'evolució no es manifesta obertament fins a l'adveniment dels éssers humans. Tot un ventall d'efectes de retard es focalitzen en l'aparició de la parla humana. Compartint les conclusions de Jos Verhulst, la posibilitat de la parla es revela com la gran fita evolutiva. En el marc d'una macroevolució ascendent la força creadora del Verb arriba a manifestar-se en la materia animada i autoconscient. Sols les sinèrgies d'un cos físic humà sense especialitzar ni animalitzar possibiliten aquest fet evolutiu trascendental.
Escrit per xavianthropus en temps de confinament, Març i Abril de l'any 2020
BIBLIOGRAFIA
L'autoconsciència, Jordi Martí i Monllau. Blog Filosofia i vida, 2018:
https://stoacatalana.wordpress.com/2018/08/26/lautoconsciencia/
L'evolució del cervell humà, Aida Gomez Robles i Chet C. Sherwood. Mètode 89, 2016. Universitat de València:
https://metode.cat/revistes-metode/monografics/levolucio-cervell-huma.html
Heterocronía, generadora de cambios biológicos, Mariana B. J. Picasso. División Paleontología Vertebrados, Museo de la Plata. Vol. 3, nº 21, Nov 2007, págs. 27-31.
http://sedici.unlp.edu.ar/bitstream/handle/10915/47291/Documento_completo__.pdf?sequence=1&isAllowed=y
Evo-devo - Biología evolutiva dels desarrollo. Laura Nuño de la Rosa, Konrad Lorenz Institute for Evolution and Cognition Research. Instituto de Filosofía Universidad Austral, 2015.
http://dia.austral.edu.ar/Evo-devo_-_Biolog%C3%ADa_evolutiva_del_desarrollo
Developmental Dynamics in Humans and Other Primates, Jos Verhulst. Verlag Freies Geistesleben, Stuttgart 1999. Adonis Press, New York 2003.
La Variedad de la Vida, Colin Tudge. Editorial Crítica, Barcelona 2001. The variety of life, Oxford University Press, 2000.
Curso de Zoología, Eugen Kolisko, Stuttgart 1932. Kolisko Archive Publications, Bournemouth 1978-1986. Editorial Rudolf Steiner, Madrid 2009.
Hombre y Animal, Hermann Popelbaum. Philosophisch-Anthroposophischer Verlag, Dornach 1975. Ed. Pau de Damasc, Cuadernos de Aproximación a la Naturaleza nº 4, Barcelona 1993.
Cosmos y Gea, Fundamentos de una nueva teoría de la evolución, Francesc Fígols. Editorial Kairós, Barcelona 2007.

I finalment tenim un altre tipus de mamífers en els que observem una hipertròfia clara de les dents canines, els ullals! Parlem, és clar, dels carnívors, com ara un gos o un gat domèstics, per exemple, o d'espècies salvatges com el llop i el tigre.
 |
| Crani de Canis lupus familiaris. Destaquen les dents canines característiques dels carnívors. |
 |
| Crani de Felis silvestris catus. Els fèlids tenen ullals ben desenvolupats i un musell curt. |
 |
| Panthera leo i Panthera tigris, els dos felins vivents més grans. Esquerra, lleó. Font Viquipèdia, Kevin Pluck CC BY 2.0 Dreta, tigre. Font Viquipèdia, ArtMechanic CC BY-SA 3.0 |
Tradicionalment s'associa el lleó amb el coratge i la valentia, simbolitzats pel seu cor (recordeu en Ricard Cor de Lleó, valent combatent, lider guerrer i rei d'Anglaterra). El cor, a la fi, pertany a la part central del cos i equilibra amb les seves pulsacions rítmiques els extrems cefàlic i abdominal. Cor i pulmons es coordinen dins del sistema sanguini per regular els ritmes cardio-respiratòris. Amb el ritme dels batecs del cor i de les respiracions, la regió toràcica armonitza les polaritats en un organisme sa. Quan el cor perd el ritme es presenta l'arrítmia, un transtorn cardíac que pot arribar a ser greu.
INESPECIALITZACIÓ HUMANA
En els mamífers i els humans la regió toràcica està clarament separada de la regió abdominal pel múscul del diafragma. Parlem així d'una frontera objectiva, que no subjectiva, entre regió abdominal i regió toràcica. La regió cefàlica, el cap, resta unida i alhora separada del tòrax pel coll. Ens trobem doncs, davant d'una clara tripartició estructural en tres grans àrees orgàniques d'influència. En els animals, les fortes descompensacions unilaterals respecte al desenvolupament relatiu de les àrees cefàlica, tòracica i abdominal queden reflectides en la seva dentició o bec. A grans trets, podem distingir en els vertebrats superiors (ocells i mamífers), i sense menystenir tots els casos intermedis menys especialitzats, tres grans impulsos o tipus d'especialització a partir de les seves dents o becs. Així, en els ocells i els mamífers rosegadors predominarien les forçes neuro-cefàliques, amb desenvolupament de becs o dents incissives. En els mamífers remugants i altres grans herbívors les forces digestives i abdominals, amb els premolars i molars ben desenvolupats. I en els carnívors les forces toràciques cardio-respiratòries, tots ells amb bones dents intermèdies o canines i amb els fèlids com a punta de llança. El lleó, la vaca i l'àliga serien els animals més representatius de cada grup.
Arribats a aquest punt, afegir que, en els humans, trobem també un ésser tripartit però equil.librat, mancat de l'especialització animal i, per tant, "lliure" gràcies a la seva indeterminació, polivalència i totipotencialitat. Estariem davant d'un ésser equilibrat i polivalent, no especialitzat. No animalitzat a la fi.
ROSTRE HUMÀ I MUSELL ANIMAL
Els humans tenim, en comparació amb els animals, una cavitat oral excepcionalment curta. No desenvolupem un musell o boca prominent com els mamífers o els rèptils, o un bec com les aus i les tortugues. Totes les etapes fetals del mamífers presenten boques curtes, però en els humans aquesta característica és manté sempre, mentre que en els mamífers es va perdent poc o molt durant el creixement i maduració.
El rostre ample i aplanat és un altre fenòmen de fetalització o retardació propi dels humans. En els primats inferiors ja trobem la tendència a reduïr el musell, amb un paladar un 30 % més curt que el dels mamífers insectívors. En els micos la reducció augmenta de mitjana fins al 40 % i en els humans fins al 65 %. En el cas de l'arc dental succeeix quelcom de similar. En els primats, respecte dels insectívors i micos inferiors, la seva longitud minva un 25 % i en humans un 65 %. Aquesta tendència cap a la reducció de l'arc dental va acompanyada de una reducció en el nombre de dents. Evolutivament la quantitat de dents en els mamífers tendeix a disminuir en general, tret d'algunes especialitzacions concretes. Mamífers antics prehistòrics mostren al voltant de 60 dents i els rèptils encara més. En simis i humans s'ha reduït significativament la longitud de la fila dental dels molars i pre-molars. En humans adults les dents són trenta-dos: 4 incissius, 2 canins, 4 premolars i 6 molars (comptant els queixals del seny, de tardana aparició) a cada mandíbula. En la dentició de llet falten els molars.
Quant a la forma de l'arc dental humà és molt diferent de la dels micos adults. En els primers es manté sempre curva, però en els segons arriba a ser casi rectangular. Primats inferiors com el tarser filipí i petits simis com els gibons mantenen però tota la vida un rostre més aplanat i humanoide.
En canvi, els simis antropomorfes han abandonat les característiques primitives i prototípiques d'un crani facial retingut, sense musell, sobretot els mascles adults que desenvolupen ullals espectaculars. Les femelles en canvi mantenen més les característiques fetals i juvenils amb unes dents més contingudes i un arc dental menys rectangular.
En l'evolució ascendent d'un arquetipus homínid moderat i retingut sembla que la questió de gènere és important. Els mascles cauen més en l'animalització i atenent al seu desenvolupament ontogènic individual podriem dir que la seva involució és notoriament major. Si un goril.la (educat amb una tutora humana) va ser capaç d'aprendre a comunicar-se amb els humans mitjançant llenguatge de signes, aquesta va ser Koko, un goril.la femella, esclar. Va arribar a dominar fins a 1000 paraules en el llenguatge de signes americà (ASL) i a comprende fins a 2000 en anglès. Un espectacular aprenentatge cognitiu per a l'espècie dels goril.les. També hauriem d'admetre en els mascles humans un cert handicap social evolutiu de caire testosterònic que contrasta amb la plasticitat i flexibilitat adaptatives femenina i que confirmaria alló de que "la dona és el futur de l'home".
LES DENTS I LA PARLA
L'estructura de la mandíbula i les dents humanes va més enllà de la pura funció masticatòria i alimenticia. La dentició no correspon amb cap dels patrons habituals dentals dels diferents tipus de mamífers com carnívors, herbívors, rosegadors, insectívors o fins i tot omnívors com els porcs, doncs la boca humana a més de formar part de l'aparell digestiu és alhora una estructura anatòmica que possibilita la parla juntament amb la laringe, l'òrgan propi de la fonació.
 |
| Esquerra, boca i dentadura humana. Font: Piqsels, imatge CC0 Dreta, meitat de l'arc dental inferior vist des de dalt. Font: Viquipèdia, Gray's Anatomy (1918), domini públic |
Degut a la uniformitat d'alçada i amplada de totes les dents i sobre tot a l'orientació vertical dels incisius i la falta d'espais entre les dents frontals, la dentadura humana forma davant de la cavitat oral una tanca continua que és essencial per a la formació de consonants.
DESENVOLUPAMENT DEL CRANI
Una cosa semblant succeeix amb el desenvolupament de la resta del cap. El crani d'un mico neonat s'assembla molt en proporció i forma al d'un humà. Simis i humans encara mantenen aquesta semblança en les etapes juvenils del desenvolupament, però en les fases posteriors solament els humans són capaços de mantenir les proporcions d'aquesta forma cranial comú inicial. El bonobo o ximpanzé pigmeu, per alguns autors una variació neotènica del ximpanzé comú, manté en la fase adulta una menor desviació del patrò de crani humanoide que l'altre ximpanzé, i també mostra un menor desplaçament posterior del foramen magnum. Es a dir, és manté relativament més proper al tipus universal humanoide en desviar-se menys de la seva figura juvenil.
 |
| Ontogènia comparada d'humans (a, b) i ximpanzés (c, d). Crani fetal i d'adult. Spotorno 1993 i 2014. Font: https://www.researchgate.net/figure/Figura-2-Ontogenia-comparada-de-humanos-y-chimpances-Extremo-superior-a-craneo-humano_fig1_264725820 |
En els micos, la forma original del crani fetal i juvenil canvia de més esfèrica a allargada, degut al creixement posterior de les mandíbules. En canvi, el neurocràni deixa de crèixer ben aviat, de manera que el creixement de la cavitat cerebral s'atura deseguida mentre continua el creixement de l'aparell masticador.
En els humans el crani tarda en ossificar-se definitivament. Neixem amb fontane.les (espais tous) i separacions entre el ossos cràneals. Aixó permet la superposició d'aquests ossos durant el naixement, degut a un crani massa gran a l'hora de passar pel canal del part. Les fontanel.les resten sense tancar després de néixer encara durant un cert temps (uns 18 mesos tarda en ossificar la fontanela anterior o major). Els ossos cranials que envolten al cervell continuen creixent fins els sis anys, amb el 90 % assolit del desenvolupament, i la resta en arribar a la pubertat. En canvi, les sutures fibroses entre els ossos del crani no ossifiquen fins a edat avançada i en alguns individus mai, com és el cas de la sutura frontal o metòpica. El desenvolupament del crani facial també s'allarga en comparació al del neurocrani. Al sis anys ha arribat al 80 % del seu creixement, però continua lentament en la pubertat i més enllà.
L'ASSUMPTE DEL MENTÓ
La mandíbula, propiament dita, o mandíbula inferior, queix o maixella, no presenta ossos premaxil.lars com la mandíbula superior. Està formada per la unió de les dos hemimandíbules o hemiarcades. En els infants humans els dos maxil.lars inferiors es mostren separats durants un temps per la símfisi o sutura mandibular, però es fusionen entre el primer i segon any de vida mitjançant l'ossificació dels teixits connectius de la símfisi. En molts altres animals pot romandre sempre sense ossificar, com per exemple en gossos i vaques, ambdós amb fort creixement frontal del crani facial.
També obsevem un desfase entre un major creixement frontal de la mandíbula i un creixement menor de l'arc dental humà. Això dóna origen a l'aparició del mentó o barbeta, un tret exclusiu nostre que no compartim amb cap animal. La mandíbula humana continua creixent una mica durant el segon septeni, quan el creixement de l'arc dental ja s'ha aturat abans coincidint amb el canvi de la dentició de llet. D'aquí el mentó lleugerament prominent.
En els simis antropoides això no passa, ja que l'arc dental continua creixent a un ritme major que la base mandibular, que també creix però menys i per tant queda sempre una mica endarrerida i fonedissa. En els simis i els altres mamífers observem també un desfase temporal en el patró de creixement de la mandíbula i de l'arc dental, però en sentit totalment contrari.
EL MISTERI DE L'OSSA INCISIVA
Els vertebrats, des dels peixos ossis fins els ocells amb bec o els mamífers amb morro, tenen dos petits ossos situats al davant de la mandíbula superior, són els premaxil.lars i són part i causa important de la prominència del musell. Envolten l'obertura nasal i part del pont del nas i baixen fins a la boca, on conformen l'àrea frontal de l'arc dental superior i allotjen les dents incissives, d'aquí el nom d'ossa incisiva. En el musell dels mamífers trobem aquests ossos separant els dos maxil.lars superiors, tot impedint-los soldar entre si els dos extrems frontals i en consequència sense cap fre que aturi el seu creixement cap endavant.
Abans de les investigacions de Goethe (1780) es creia que l'ossa incisiva era inexistent en humans. Contrariat amb aquesta afirmació, Goethe va descobrir finalment en alguns cranis humans restes de la sutura entre el maxil.lar i l'os incisivum, fet que confirmava l'existència dels premaxil.lars en humans. Aquesta sutura es pot trobar en una quarta part dels cranis d'adults humans i en la majoria dels juvenils. Sempre es troba a la banda del paladar, formant un cercle tancat per darrera dels incisius, i mai a la part externa del crani facial, on queda ja engolida pels maxil.lars al final de l'etapa embrionària.
La falta de musell en els humans, per tant, va associada a aquest ràpid engoliment dels ossos premaxil.lars per part dels maxil.lars superiors durant el desenvolupamernt primerenc, cap al final del tercer mes de gestació.
Goethe ja va veure en aquesta ràpida incorporació dels premaxil.lars una expressió de la resistència humana al desenvolupament del musell. Considerava que un dels propòsits d'aquesta resistència era la preservació d'una fila de dents tancada (detall important per a la parla). Molts mamífers presenten diastemes o espais entre les dents, com ara els remugants i els rosegadors entre els molars i els incisius, i els primats també mostren uns espais per a acomodar les parts superiors de cadascun dels grans ullals en el maxil.lar contrari pertinent. En els infants humans trobem també habitualment aquests anomenats "espais primat" i altres diastemes de creixement en la dentició temporal o de llet, un fet que facilita l'encabiment de les futures peces dentàries definitives de major tamany.
Respecte a la sutura incisiva, a més de vestigis en el paladar, en els grans simis trobem també restes d'ella a la banda exterior del crani, ni que sigui temporalment. Bolk va estudiar el tema en 1912 i va trobar que en els ximpanzés la sutura externa pot desaparèixer ja abans de néixer, en els orangutans es perd durant l'infància i més tard en els gori.les. En canvi perduren en els adults de les mones aranya.
La tendència a l'escurçament de l'os incisivum i del musell, tot mantenint un cap arrodonit, ja la trobem en primats primitius com els tarsers (amb fòssils des de l'Eocé, a principis del període Terciari), encara que hi ha importants retrocessos en etapes més tardanes de la evolució dels primats, com en els papions, gelades, drils i mandrils (tots ells amb un musell força prominent i ullals impressionants en els mascles). Aquesta tendència cap a la supressió del musell es manté però com a característica evolutiva del primats, i en els simis superiors s'observa clarament un tancament ràpid del premaxilars, encara que no aconsegueixen frenar del tot el desenvolupament del musell. Solament els humans assolim la supressió completa, obrint el camí cap a la parla. Un camí fressat de confluències sinèrgiques com les descrites fins aquí i d'altres no comentades encara.
LA COMPOSICIÓ SINÈRGICA
Altres característiques humanes són, per exemple, l'independència del ritme respiratori, al no estar condicionat per una locomoció quadrúpeda (una respiració per cada pas o gambada, o bé la independència respiratòria respecte de la termoregulació per ventilació pulmonar (el panteix dels gossos). Amdós circumstàncies faciliten la parla. També la llengua que surt des de més avall i enrera, formant la paret anterior mòbil de la faringe (molt important per modular els sons) i una laringe o garganxó situada en els adults humans molt més avall que en els mamífers (podem respirar independentment per la boca o el nas), i en angle recte respecte de la cavitat bucal (aquesta posició permet un inventari molt ample de sons). Les cordes o plecs vocals, a més, són de constitució muscular en els humans i membranoses en els primats inferiors, mentre que en els simis antropoides són d'una constitució intermèdia.
Al cap i a la fi tot un ventall de processos sinèrgics convergents per a possibilitar la parla humana.
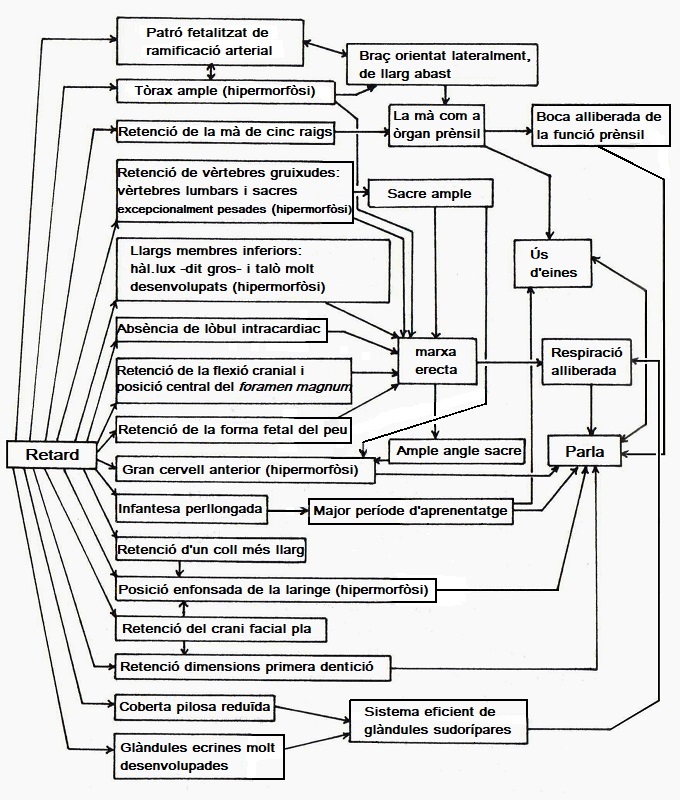 |
| Diagrama de les interconnexions entre les diferents consequències de la paidomorfòsi en els éssers humans (Jos Verhulst, 1999). |
Les diferents consequències del retard generalitzat en el desenvolupament humà respecte de la forma prototípica mamífera o primat, i fins i tot vertebrada o cordada, es junten armoniosament en el cos humà tot formant una composició sinèrgica, un conjunt potencialment molt superior a la simple suma de les seves parts.
L'objectiu del retard constant i creixent dels primats al llarg de l'evolució no es manifesta obertament fins a l'adveniment dels éssers humans. Tot un ventall d'efectes de retard es focalitzen en l'aparició de la parla humana. Compartint les conclusions de Jos Verhulst, la posibilitat de la parla es revela com la gran fita evolutiva. En el marc d'una macroevolució ascendent la força creadora del Verb arriba a manifestar-se en la materia animada i autoconscient. Sols les sinèrgies d'un cos físic humà sense especialitzar ni animalitzar possibiliten aquest fet evolutiu trascendental.
Escrit per xavianthropus en temps de confinament, Març i Abril de l'any 2020
BIBLIOGRAFIA
L'autoconsciència, Jordi Martí i Monllau. Blog Filosofia i vida, 2018:
https://stoacatalana.wordpress.com/2018/08/26/lautoconsciencia/
L'evolució del cervell humà, Aida Gomez Robles i Chet C. Sherwood. Mètode 89, 2016. Universitat de València:
https://metode.cat/revistes-metode/monografics/levolucio-cervell-huma.html
Heterocronía, generadora de cambios biológicos, Mariana B. J. Picasso. División Paleontología Vertebrados, Museo de la Plata. Vol. 3, nº 21, Nov 2007, págs. 27-31.
http://sedici.unlp.edu.ar/bitstream/handle/10915/47291/Documento_completo__.pdf?sequence=1&isAllowed=y
Evo-devo - Biología evolutiva dels desarrollo. Laura Nuño de la Rosa, Konrad Lorenz Institute for Evolution and Cognition Research. Instituto de Filosofía Universidad Austral, 2015.
http://dia.austral.edu.ar/Evo-devo_-_Biolog%C3%ADa_evolutiva_del_desarrollo
Developmental Dynamics in Humans and Other Primates, Jos Verhulst. Verlag Freies Geistesleben, Stuttgart 1999. Adonis Press, New York 2003.
La Variedad de la Vida, Colin Tudge. Editorial Crítica, Barcelona 2001. The variety of life, Oxford University Press, 2000.
Curso de Zoología, Eugen Kolisko, Stuttgart 1932. Kolisko Archive Publications, Bournemouth 1978-1986. Editorial Rudolf Steiner, Madrid 2009.
Hombre y Animal, Hermann Popelbaum. Philosophisch-Anthroposophischer Verlag, Dornach 1975. Ed. Pau de Damasc, Cuadernos de Aproximación a la Naturaleza nº 4, Barcelona 1993.
Cosmos y Gea, Fundamentos de una nueva teoría de la evolución, Francesc Fígols. Editorial Kairós, Barcelona 2007.